
Conocimiento de la biodiversidad: El programa Ecosistemas y Recursos Naturales, de acuerdo con la misión institucional, continuó durante la vigencia 2019-2022 aportando al Inventario Nacional de la Biodiversidad, mediante los inventarios biológicos, el crecimiento y fortalecimiento de las colecciones y la divulgación de información. Así, durante esta vigencia el grupo de flora, fauna, recursos hidrobiológicos, recursos genéticos y biología del suelo exploramos 31 localidades de la Amazonía colombiana (Fig. 1), publicamos 133 contribuciones relacionadas con la diversidad biológica (Anexo 1.), incorporamos 32.235 ejemplares a las colecciones, 1.269 taxones nuevos para las colecciones (Fig. 2), 364 registros nuevos para el país y 34 especies nuevas para la ciencia. En la vigencia, a partir de los inventarios de biodiversidad en las nuevas localidades se ha ampliado la distribución geográfica de un número importante de especies amenazadas y endémicas, 44 amenazadas, 84 endémicas y 334 en apéndices CITES.
Las cifras acá reportadas están soportan en ejemplares voucher debidamente curados en las colecciones con la ayuda de varios especialistas invitados y dispuestas para consulta en línea; las publicaciones indican un intenso trabajo colaborativo con varios grupos de investigadores nacionales y de diversos países. De igual forma, y atendiendo las necesidades de información regional, se hizo un esfuerzo considerable en divulgar los resultados para los usuarios en la Amazonia, tanto comunidades como tomadores de decisión.
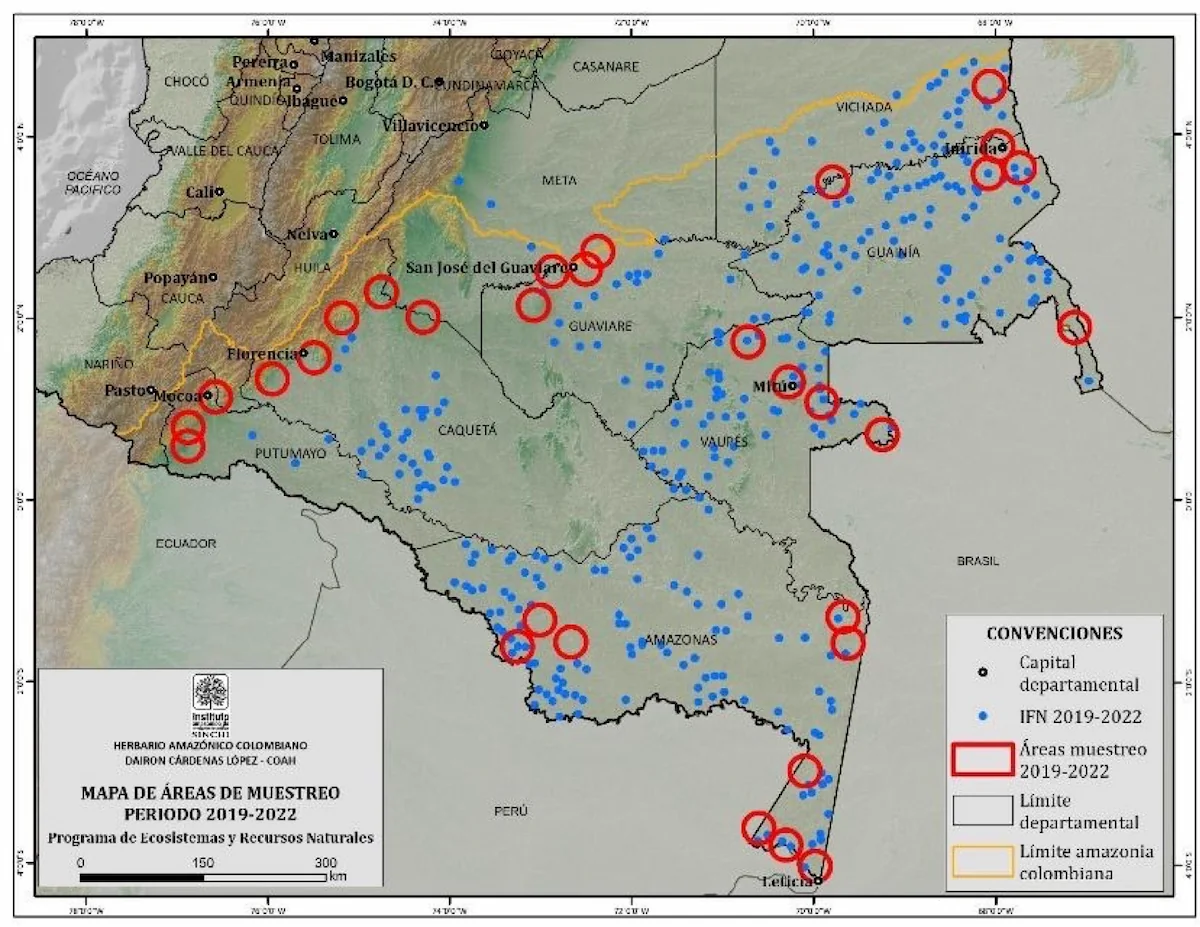
Figura 1. Localidades de estudio del programa Ecosistemas y Recursos Naturales 2019-2022
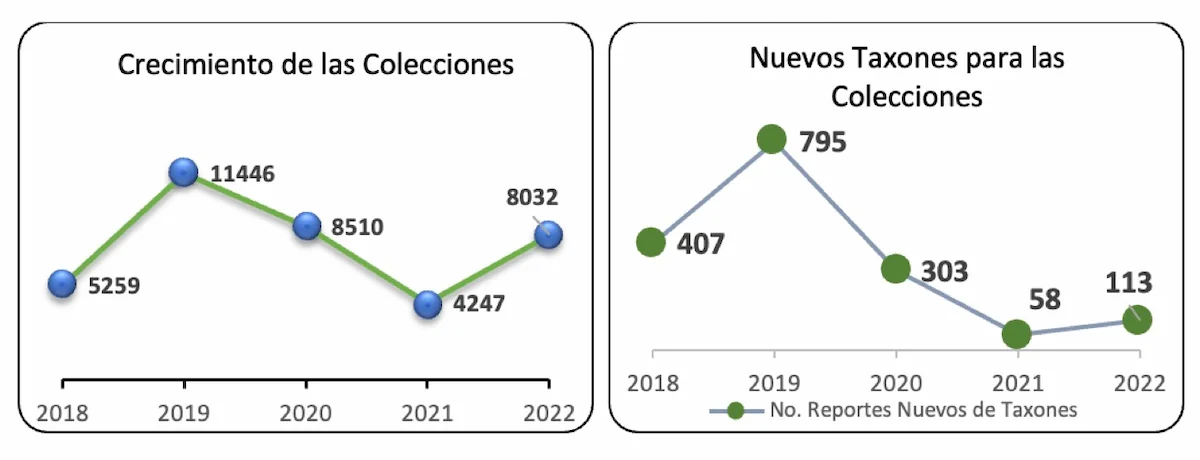
Figura 2. Ejemplares y nuevos taxones incorporados a las colecciones del Instituto Sinchi 2019-2022
Uso, manejo y conservación de la biodiversidad
Flora
Se avanzó en el conocimiento del uso y conservación de la flora Amazónica en varios frentes, principalmente enfocado en: i) la oferta y dinámica de las especies maderables y no maderables, ii) de las especies invasoras, iii) de las especies amenazadas y iv) la funcionalidad y dinámica de los bosques Amazónicos y v) La valoración económica de los bosques
Resultados
i) Nos hemos enfocado en el monitoreo del crecimiento de siete especies maderables ampliamente usadas en la región amazónica colombiana, así: abarco amazónico (Cariniana domestica), cedro (Cedrela odorata), achapo (Cedrelinga cateniformis), palo de arco (Handroanthus serratifolius), quinilla (Manilkara bidentata), acapú (Minquartia guianensis) y macacauba ó granadillo (Platymiscium stipulare). Este trabajo ha permitido generar información que aporta al manejo sostenible de las especies. Se resalta que Cariniana domestica presentó el mayor valor promedio de crecimiento diamétrico con 1,76 cm/año, seguido de Cedrelinga cateniformis con 1,69 cm/año y por Cedrela odorata con 1,2 cm/año. La mortalidad para este grupo de especies varió entre el 50 y el 9% de sus individuos, con un promedio de 14.8%. Esta información es de gran importancia para los Planes de Manejo que sustentan el aprovechamiento sostenible de estas especies en la región Amazónica colombiana, con base en información del comportamiento de cada una de ellas. A su vez, se ha generado información sobre la abundancia y dinámica de cinco especies no maderables usadas por comunidades indígenas en el departamento del Amazonas; tres especies usadas en cestería: Yaré (Heteropsis, Araceae), Bejuco-Boa (Desmoncus polyacanthos, Arecaceae), Guarumo (Ischnosiphon arouma Marantaceae); y dos especies usadas en medicina tradicional: Chuchuwasa (Maytenus sp. – Olacaceae) y Huacapurana (Campsiandra comosa Caesalpiniaceae).
ii) Las invasiones biológicas son unos de los grandes problemas de conservación que sufren las áreas naturales en todo el mundo. A partir del trabajo interinstitucional y para el caso colombiano la información del Herbario Amazónico Colombiano se logró generar una base de datos mundial sobre las especies de invasoras (van Kleunen et al 2019) , en la se provee información sobre la región de origen, la región donde es reportada como invasora; También proporcionamos varias variables que se pueden utilizar para filtrar los datos de acuerdo con la calidad y la integridad de las listas de taxones exóticos, que varían entre las combinaciones de regiones y fuentes de datos. La base de datos abierta [1] permite que la información sobre especies invasoras está disponible para todo público y que la identificación de invasiones biológicas sea a nivel global.
iii) La generación de información sobre especies amenazadas se ha concentrado por un lado en la identificación y caracterización del estado de amenaza de las especies (Cárdenas et al. 2019b, López et al 2019), la identificación de nuevas especies (Barona-Colmenares & Hágsater. 2020), la categorización de grupos de especies como los helechos arborescentes (Cárdenas et al 2019d), en donde a partir de la evaluación del estado de poblaciones naturales y siguiendo los protocolos de la IUCN se categorizaron 140 especies de las familias Cyatheaceae y Dicksoniaceae y se documentaron 19 nuevos registros para Colombia; se encontró que 46 especies de helechos arborescentes están en alguna categoría de amenaza: 18 En Peligro Crítico, 14 En Peligro y 14 en Vulnerable (Cárdenas et al 2019d). Por otro lado, hemos recaudado de información paleobiológica de la flora tropical y se reafirma que la palo-ecología es clave para entender los bosques modernos (Carvalho et al. 2021, Heijink et al 2020, Serrano et al 2021). Frente a las especies amenazadas también se ha avanzado en la implementación de los planes de manejo de Cedro, Vainillas y Zamias, en donde en todos los casos se han identificado nuevas poblaciones naturales y se han realizado procesos de propagación asistida y para el caso de Cedro (Cedrela odorata) se ha generado información sobre la variación genética de sus poblaciones naturales.
iv) A partir del trabajo interinstitucional (Davies et al 2021) se ha generado una gran cantidad de información relevante sobre la distribución de especies de la Pan-Amazonia (ter Steege et al 2019), su relación con los síndromes de dispersión (Correa et al 2022), sobre como las condiciones hidrológicas locales modulan la composición de los bosques tropicales y como al tener en cuenta la topografía y el gradiente hidrológico local se explica mucho mejor las variaciones en diversidad y de respuesta de los bosques a lo largo de la pan-Amazonía (Marca-Zeballos et al 2022). Así como refinar los cálculos de la biomasa que se almacena en los bosques tropicales (Cushman et al. 2021) y como la variación demográfica de las especies altera profundamente los flujos de carbono a lo largo de los gradientes latitudinales entre los trópicos y las zonas templadas (Needham et al 2022).
Estos trabajos mancomunados, han permitido avanzar en el conocimiento sobre la dinámica del carbono y su relación con los grandes árboles en diferentes tipos de bosques (Piponiot et al 2022); así como afinar los modelos cuantitativos de predicción de especies que se podrían encontrar en la Amazonía (ter Steege et al 2020, Pos et al 2022). Finalmente es de destacar que hemos avanzado en el conocimiento de los factores, causas y condiciones de la mortalidad de los árboles y como los daños de ramas y tronco de los árboles aumenta el riesgo de mortalidad del árbol (Zuleta et al 2022a) y como algunos rasgos funcionales están relacionados con la topografía (Zuleta et al 2022b).
Finalmente, a través del monitoreo de la Red de Parcelas Permanentes de 1 hectárea del Instituto SINCHI (34 parcelas en total), se han identificado las afectaciones del cambio climático sobre los bosques naturales. Se obtuvo una tasa de reclutamiento anual promedio de 1,5 % ind/ha/año con variaciones entre 3,1 y 0,3 % ind/ha/año. La tasa de mortalidad anual presentó un promedio de 1,81% /ha/año con un valor máximo de 3,72% /ha/año y un mínimo de 0,34 % /ha/año. La tasa de crecimiento anual (relación entre crecimiento diamétrico más biomasa de reclutas, menos biomasa de árboles muertos) presentó una variación con valores negativos de hasta -4,0 Ton/ha/año y un máximo de 5,23 Ton/ha/año. Este monitoreo igualmente ha permitido generar alertas tempranas frente a grupos de especies que están siendo afectadas más severamente por el cambio climático, así como identificar especies más resilientes.
v) La valoración económica de los bosques ha permitido calcular el costo de perder una hectárea de bosque, así como identificar la valoración no monetaria de los bosques por parte de sus usuarios más directos: los pueblos indígenas.
Recientemente se generó un documento que condensa el estado del conocimiento de las especies útiles de la Amazonía colombiana (Cárdenas et al. 2022) en donde se identifican 2286 especies útiles en la región.
Recomendaciones Flora
-Continuar con los estudios a largo plazo de la dinámica de la estructura y composición de los bosques de la Amazonía colombiana. La generación de información sobre las respuestas de las plantas al cambio climático permitirá generar alertas tempranas y un mejor entendimiento de cómo se estructuran y reaccionan los bosques Amazónicos
-Avanzar en el conocimiento de fenológico y de requerimientos de propagación de especies de plantas poco conocidas.
-Enfocar esfuerzos para la generación de información sobre la funcionalidad de las especies
Fauna
Los habitantes amazónicos, especialmente las comunidades indígenas tienen una estrecha relación con las especies silvestres que se encuentran en sus territorios, muchas de ellas fundamentales en su seguridad alimentaria y con las cuales mantienen unos vínculos espirituales que derivan en una serie de normas culturales para su uso. Este uso, cuando ha sido posible mantenerlo bajo un manejo tradicional, se ajusta normalmente a cantidades estrictas para el consumo y para compartir comunitariamente las presas. No obstante, fenómenos como el crecimiento de las ciudades amazónicas y el desplazamiento hacia las cabeceras municipales de las familias por diversos servicios, hace necesario contar con recursos económicos para mantenerse en las ciudades y en este sentido, las presas se convierten en una transacción segura para contar con dicho recurso.
Desde el grupo de fauna hemos buscado entender cuáles son las especies que están implicadas en la dieta de las comunidades, cuál es la magnitud del uso, la oferta natural y las tendencias de las presiones a la que están sometidas; esto con la finalidad de implementar, si se requiere, manejos ajustados al uso responsable, que garanticen la sostenibilidad y la conservación de las especies. Se aborda desde el monitoreo comunitario, investigación que tiene sentido para las comunidades locales, tiene un valor práctico para su bienestar y construye una comunidad del aprendizaje que reconoce que cada integrante posee conocimientos y recursos únicos que pueden contribuir a la gestión (Moller et al. 2009, Robson et al. 2009).
Los métodos empleados constituyen aproximaciones de corto plazo en las que se identifican especies a monitorear al establecer una presunta presión a partir de la percepción de abundancia y la frecuencia de uso obtenida mediante entrevistas (Weller 2007, Van Holt et al. 2011, Bogartti & Halgin 2011); y una de largo plazo en la cual las comunidades hacen un seguimiento sistemático del uso y disponibilidad de la fauna mediante metodologías clásicas para evaluar abundancias poblacionales (Buckland et al. 2001, Rowcliffe et al. 2008, Townsend 1996, Bodmer 1994, Redford & Robinson 1987)
Resultados
Se evaluaron los insectos útiles para el consumo en el departamento del Vaupés a raíz de un primer trabajo que se adelantó al sur del departamento en el año 2013 (Osorno et al. 2014). A pesar de la poca información disponible se hizo una revisión general para el país y se consolidó un total de 69 especies de insectos comestibles reportados, distribuidos en 49 géneros, 27 familias y 10 órdenes, que son consumidos en aproximadamente 13 grupos étnicos pertenecientes principalmente a las regiones Amazónica y Caribe (Gasca-Alvarez & Costa-Neto 2021). En las comunidades de Santa María de Itapinima y Piracemo, municipio Mitú, se registraron 14 especies comestibles de insectos pertenecientes a 10 familias y 5 órdenes, Coleoptera, Blattodea, Hymenoptera, Lepidoptera y Hemiptera; las más frecuentes e importantes en la dieta son el mojojoy Rhynchophorus palmarum, la manivara Syntermes spinosus, la hormiga arriera Atta laevigata y varias especies de orugas de polillas llamadas tapurú como Lusura chera (Lepidoptera:Notodontidae) y Polybia liliacea (Hymenoptera: Vespidae); se indagó por el calendario ecológico de estas especies, técnicas de cosecha y sus formas de preparación y consumo tradicional ( 2021, 2022).
Se avanzó en el seguimiento, durante 15 meses (septiembre 2017- diciembre 2018), de la comercialización de animales silvestres para el consumo en la plaza de San Victorino en Mitú. Se registraron 353 eventos de comercialización equivalentes a 2.035 Kg, provenientes de 72 comunidades y que incluyeron 9 taxones de los cuales la lapa fue la especie más comercializada, como es habitual en la Amazonia colombiana. En 5 comunidades indígenas, con niveles de comercialización contrastantes se exploró la relación entre comercialización, percepción de abundancia y acatamiento a las normas ancestrales. Se encontró que de 92 especies o grupos de especies de fauna que hacen parte de la alimentación de estas comunidades, la lapa (Cuniculus paca), la guara (Dasyprocta fuliginosa), el tintín (Myoprocta pratti), la pava (Penelope jacquacu) y la danta (Tapirus terrestris) registran los mayores valores de relevancia en el dominio cultural. A la guara, que presenta bajos niveles de comercialización, pero alta relevancia cultural, la mayoría de informantes la consideran abundante mientras a la danta y al cafuche (Tayassu pecari) los perciben como especies raras. Por efecto de la comercialización, la guara parece estar reemplazando a la lapa en la alimentación de las comunidades indígenas. La información generada en esta investigación se utilizó para un diálogo constructivo con las autoridades indígenas y la institucionalidad en Mitú, con el fin de llegar a acuerdos internos para el uso sostenible de la lapa y de otras especies de cacería comprometidas en la comercialización, con el fin de que las comunidades no sacrifiquen la alimentación por este tipo de transacciones económicas. Actualmente se llevan a cabo conversaciones con los cazadores e intermediarios de esta comercialización al interior de cada comunidad.
Desde 2018 el Instituto Sinchi, con el apoyo del proyecto Gef Corazón de la Amazonia, avanza en conjunto con las comunidades indígenas de la Estrella Fluvial Inírida en la evaluación de la fauna de consumo para su manejo. El monitoreo ha identificado a la fecha 69 especies de fauna usadas para el consumo; así como la magnitud de su aprovechamiento tanto para consumo como para comercialización. Se acumulan 35 meses de seguimiento, 2.431 presas capturadas, 2.137 faenas reportadas por 495 cazadores en 24 comunidades, con un aprovechamiento promedio 833 presas/año y una biomasa extraída de 6.346 kg/año. Actualmente se evalúa la disponibilidad de sus poblaciones a partir de fototrampeo‚ estimación con transectos lineales y captura por unidad de esfuerzo; así como la sostenibilidad del uso‚ para proponer medidas de manejo que garanticen su conservación y la seguridad alimentaria de las comunidades de la EFI. En el proyecto han participado 112 monitores de 24 comunidades y seis etnias (curripaco, puinave, sikuani, piapoco, cubeo y tukano), hoy el grupo se consolida con 30 monitores. La estrategia de trabajo se fundamenta en el fortalecimiento continuo de capacidades y la divulgación periódica de resultados en un lenguaje adecuado‚ de tal forma que los monitores pueden discutir los resultados con sus comunidades. Con los resultados hasta la fecha ya se han propuesto medidas de conservación sobre áreas y sobre especies; además de medidas de gobernanza.
Los datos acopiados por el Instituto, tanto en monitoreos comunitarios de largo plazo como en evaluaciones rápidas por consenso cultural, fueron analizados para establecer la diversidad de fauna usada para el consumo, la variación de la dieta en amplitud y composición, y la influencia de algunos factores socioambientales en las especies de fauna consumidas. Registramos 124 especies de fauna silvestre usadas para consumo en la Amazonia colombiana (45 especies de aves, 38 de mamíferos, 17 de reptiles, 6 de anfibios y 18 especies de invertebrados), con amplitudes de dieta que varían entre 31 y 74 especies por localidad. Tal como se registra en otros trabajos (Stafford et al. 2017), se encontraron similitudes en la estructura de la dieta entre las comunidades de sectores geográficos cercanos. En cuanto a la composición, los mamíferos son el grupo que más aporta en biomasa en todas las localidades estudiadas, seguidos de los reptiles. La diversidad de especies de fauna en la dieta parece estar relacionada con su disponibilidad a nivel local, la tasa de retorno de las presas cazadas y el mantenimiento de manejos tradicionales. Estos análisis y comparaciones a nivel regional permiten inferir que los cambios socioculturales que han sufrido las comunidades indígenas han impactado en la diversidad de la fauna incluida en la alimentación, al parecer con efectos en la sostenibilidad del uso por lo cual se hace necesario continuar identificando e implementando propuestas de manejo culturalmente viables que propendan por un uso sostenible de la fauna (Atuesta-Dimian et al. 2021).
Recomendaciones Fauna
Continuar construyendo la line base de diversidad de fauna en los distintos ecosistemas amazónicos y seguir haciendo esfuerzos para asegurar la apropiación de dicha información y nuevo conocimiento por parte de los usuarios directos, las comunidades y sus autoridades en los territorios amazónicos.
Ampliar y reforzar las alianzas con grupos e investigación a nivel mundial y en la panamazonia con el fin de avanzar de manera más efectiva en la investigación sobre la diversidad amazónica y en las implicaciones del cambio climático sobre dicha diversidad.
Dada las preocupaciones de las comunidades sobre la pérdida de conocimiento tradicional para el manejo de sus recursos, es necesario idear en conjunto con ellas nuevas rutas de transmisión en las cuales el nuevo conocimiento que se deriva de los ejercicios de monitoreo comunitario de fauna silvestre pueda aportar. Por ello, es importante continuar en el fortalecimiento de capacidades de las comunidades en torno al uso y manejo de su fauna, recrear los conocimiento y normas culturales, y en este sentido fortalecer la gobernanza y proteger su seguridad y soberanía alimentaria.
Profundizar en el conocimiento sobre el uso de insectos para consumo y su sostenibilidad. Es un tópico poco explorado y de gran potencial tanto desde el punto de vista nutricional como potencial alternativa económica.
Los distintos ejercicios de valoración de la fauna de consumo en la Amazonia y las experiencias de monitoreo comunitario se dispondrán de tal manera que la información sea útil para la conformación de una red de actores que estén informados y puedan tomar decisiones en pro de su gobernanza y de la conservación de las especies, los bosques y humedales.
Recursos hidrobiológicos
El estado de los ríos, lagos y humedales se ha convertido en una prioridad principalmente porque son fuente directa de agua, biodiversidad, alimento (especialmente por peces), recreación, transporte en la Amazonia colombiana y aportes a la economía regional, (Instituto Sinchi 2013), por tanto, conocer sus características, disponibilidad y calidad de estos ambientes, la biota presente, las dinámicas de uso, el comportamiento biológico de recursos acuáticos de especial interés socioeconómico y otras dinámicas es necesario para mantener la disponibilidad y calidad de estos ambientes para los pobladores de la región.
Desde esa óptica, el grupo de Ecosistemas Acuáticos ha abordado diferentes aspectos relacionados con la caracterización de estos ambientes a partir de la biodiversidad, fisicoquímica del agua, hidrología y valoración por índices de calidad ambiental con el fin de conocer el estado y salud de los mismos. De igual manera se ha podido adelantar el monitoreo de las comunidades de algas, macroinvertebrados y peces que permitan evaluar el efecto de la deforestación en la riqueza y abundancia de estos grupos biológicos. En razón a su importancia regional, se realizan evaluaciones de aspectos biológicos y reproductivos de especies de peces de interés comercial y de autoconsumo, se establecen monitores en la comunidad para mantener un registro básico de datos que contribuyan a orientar planes de manejo pesquera y la toma de decisiones regionales y locales en la cuenca.
En la evaluación de ecosistemas acuáticos se tiene en cuenta al índice de integridad biológica (IBI) a partir de la comunidad de macroinvertebrados (Karr, 1981) y el índice de calidad de hábitat partiendo de los fundamentos establecidos por la Agencia de Protección Ambiental (EPA) y desarrollados por Barbour et al (1999). Igualmente, evaluaciones en diferentes regiones de la Amazonia –principalmente en el marco de las Expediciones BIO– proporcionan la información de línea base para documentar la biodiversidad acuática. Mientras que el trabajo sobre recursos pesqueros con fines comerciales de consumo, ornamentales y pesca deportiva, ha contado con el aporte de comunidades indígenas y campesinas en sectores de las cuencas altas de los ríos Caquetá, Putumayo y la Estrella Fluvial del Inírida. mediante el registro de información biométrica de los peces, unidades de esfuerzo de pesca, ingesta de proteína y descripción socioeconómica mediante entrevistas.
Resultados
Los estudios en ecosistemas acuáticos realizados por el Instituto Sinchi en la Amazonia colombiana se han orientado en varias líneas de investigación. Una de ellas se refiere a la caracterización física y química de los ecosistemas acuáticos, la cual permite conocer las propiedades que reflejan las condiciones biogeoquímicas y/o de contaminación de las aguas. Los ríos analizados en la vigencia 2019 – 2022 presentaron con baja mineralización, pH con tendencia a la acidez y altas concentraciones de oxígeno de la serranía de La Lindosa en Guaviare y en el área rural de Mitú (Vaupés) relacionada con las características geológicas de estas zonas. En el caso de los humedales de la zona rural de Leticia mantienen las condiciones naturales propias de estos ecosistemas amazónicos, no obstante, se observa que los humedales urbanos están altamente intervenidos, que se evidencia por los altos valores de conductividad, DBO5, DQO y en especial los coliformes. Esta información se acompaña de parámetros morfométricos de las cuencas, tales como área y perímetro de la cuenca y longitud del cauce principal, igualmente se realizaron modelos lluvia-escorrentía.
Para ecosistemas muestreados en los departamentos de Guaviare y Meta (Vistahermosa), los Índices de calidad ambiental de los humedales (Solano-Figueroa et al 2011), establecen que los cuerpos de agua analizados presentaron calidad de hábitat y de IBI en buenas condiciones, no obstante, están afectados por ecosistemas terrestres en transformación como resultado de tala de bosque para cultivos y pastos para ganadería; en campo se pudo evidenciar la reducción del bosque ribereño en la ronda hídrica de algunas quebradas; por tanto, se ubican en zonas de priorización de restauración en rondas hídricas (SIATAC, 2020).
Respecto al monitoreo que se llevan a cabo en las zonas de influencia de Florencia, Belén de los Andaquíes y otros municipios (Caquetá), Leticia (Amazonas), Mitú (Vaupés) y Puerto Leguízamo (Putumayo), principalmente en sistemas de quebradas, brindan información para comprender los efectos de la degradación de ecosistemas naturales –incluido el calentamiento global– en las comunidades de peces y macroinvertebrados. Los resultados de estos análisis demuestran que si existe un efecto del uso del suelo en la estructura de las comunidades de peces y la físico-química del agua. La mayoría de las presiones antrópicas locales sobre los cuerpos de agua (cambios en la dirección del drenaje, modificaciones de las márgenes, aportes de materia orgánica y/o nutrientes provenientes de la agricultura o asentamientos urbanos) afectan los componentes estructurales del hábitat acuático, se ha podido encontrar una evidencia de esto en la disminución de abundancias en los macroinvertebrados acuáticos monitoreados, igualmente, se notan cambios en la riqueza e incremento en la densidad del perifiton por la contaminación derivada del aporte de vertimientos de los asentamientos humanos..
En el reconocimiento de la fauna íctica presente en los ecosistemas acuáticos de la región Amazonia, se registraron 1104 especies, de estas especies, 170 son utilizadas con fines de consumo y comercio y 365 son extraídas de su medio natural para ser comercializadas como ornamentales. No obstante, para comunidades que habitan la cuenca del río Cuduyarí (Vaupés), se reconoció el consumo 77 de las 87 especies registradas para esta cuenca. Esto es solo un ejemplo de la importancia del recurso íctico en la seguridad alimentaria de los habitantes de la región. Pero a pesar de su importancia, varias de las especies utilizadas no cuentan con suficiente información biológica y pesquera para establecer su estado de conservación o amenaza, es así que se continuó el reconocimiento y seguimiento multitemporal a peces de importancia socioeconómica en algunos sectores de la región, que permitan determinar tallas de madurez sexual (punto de referencia para el manejo), épocas de reproducción, caracterización de los métodos de pesca, zonas de captura, cadenas de valor asociado al recurso y determinación de conflictos asociados. Fruto de ello, es el establecimiento de los acuerdos comunitarios en pesca para la zona Ramsar Estrella Fluvial Inírida (Resolución Aunap 2020) y avances en la historia de vida de los peces de importancia comercial y ornamental sobre cerca de 50 especies en las cuencas de los ríos Caquetá, Putumayo y en la EFI. Igualmente, se avanza con iniciativas hacia acuerdos comunitarios de pesca en sectores de la cuenca del río Caquetá (Sistema lagunar Peregrino) y cuenca del río Putumayo (Microcuenca río Caucayá).
En el contexto binacional, para los encuentros técnicos sostenidos con entidades peruanas para la pesca de consumo, se han definido las principales especies de consumo utilizadas en la Zona de Integración Fronteriza (ZIF) Perú – Colombia y se revisaron estrategias de comanejo comunitario. Se consideró necesario implementar actividades orientadas a la obtención de la información pertinente, para las 31 especies de peces que fueron priorizadas.
En el marco de la política pública y ordenación de cuencas, con el acompañamiento del Instituto Sinchi en 2021 y 2022 se culminó la formulación del POMCA del río Cuduyarí en el Vaupés, el cual había iniciado la corporación CDA desde 2016. Este trabajo contó con la activa participación de la comunidad materializada en los talleres participativos realizadas en la cuenca con representantes de 21 Comunidades Indígenas, quienes fueron sensibilizados sobre la construcción del POMCA y estuvieron comprometidos aportando sus experiencias y visiones del territorio.
Recomendaciones Recursos hidrobiológicos
La mayoría de las presiones antrópicas sobre los cuerpos de agua afectan los componentes estructurales del hábitat acuático, por lo tanto, se debe mantener un esfuerzo de caracterización de humedales para definir tipos, áreas y condiciones de estado mediante variables fisicoquímicas del agua, biológicas, hidrológicas y socioculturales, que permitan tomar medidas de gestión, teniendo en cuenta los indicadores, (indicadores de calidad fisicoquímica del agua -ICOS e ICA, indicadores de la calidad del hábitat, indicadores biológicos e Índices de Integridad Biológica), se proyecta aplicar herramientas del SIG que permitan espacializar la calidad de estos ecosistemas acuáticos. Los mapas resultantes pueden ser la base del análisis de riesgos, de la toma de decisiones y la ordenación del territorio.
Igualmente, el caracterizar y evaluar la microbiota acuática (bacterias/algas/hongos acuáticos) para el desarrollo de estrategias de biorremediación como aporte a la generación de soluciones basadas en la naturaleza, como cuantificar el efecto de los macroinvertebrados frente a la descomposición de materia orgánica como aporte en la productividad del ecosistema y en el ciclo global del carbono y realizar estudios de ADN Ambiental para ampliar los inventarios de los grupos biológicos, generan especial interés.
En esa ruta, es importante poder establecer protocolos de monitoreo participativo de la calidad de los ambientes acuáticos, por medio de pilotos en el marco del trabajo con comunidades indígenas, campesinas tanto en zonas rurales como en los cascos urbanos. Igualmente continuar con el registro de información a largo plazo para evaluar y reconocer las dinámicas de uso de comunidades biológicas de alto valor socioeconómico, en pro de favorecer y fortalecer la gobernanza y soberanía alimentaria de las comunidades locales.
En ese sentido, frente a la gran diversidad cultural del territorio, los estudios de biodiversidad acuática, de peces y en general de los humedales en la Amazonia colombiana en un contexto social o de conocimiento tradicional, hace necesario fortalecer la percepción y entendimiento acerca de estos grupos biológicos y de los humedales, por parte de la sociedad local que faciliten la implementación de estrategias de conservación y manejo del recurso. Por tanto, la información generada debe ser dispuesta mediante piezas y estrategias de comunicación adecuadas, útil para la conformación de una red de actores que estén informados y puedan tomar decisiones en pro de su gobernanza y de la conservación de las especies, los bosques y humedales.
Recursos genéticos y microorganismos
Los microorganismos (bacterias, arqueas, protozoos y hongos) constituyen un componente abundante de la biomasa total de organismos que habitan la Tierra y representan la mayor fuente de biodiversidad (Gans et al., 2005; Sogin et al., 2006). Además, desempeñan funciones críticas en procesos biogeoquímicos naturales, manteniendo el funcionamiento de los ecosistemas (Falkowski et al., 2008). Teniendo en cuenta que los microorganismos desempeñan un papel esencial en el funcionamiento del ecosistema, determinar su abundancia, composición y función, permite generar información para el establecimiento de los llamados «rangos de operación normales» para hábitats microbianos, indicando cuál es el nivel de diversidad microbiana y el funcionamiento que se espera en hábitats que funcionan «normalmente», las desviaciones de este «rango de operación normal» pueden interpretarse como perturbaciones de los ecosistemas en estudio.
Durante los años 2019 al 2022 se evaluó la composición de las comunidades microbianas en ambientes lóticos y terrestres de diferentes ecosistemas de la región Amazónica. Usando técnicas de nueva generación (NGS) como la tecnología Illumina Miseq, que permite el estudio de microorganismos no-cultivables o de difícil aislamiento, lo cual favorece la accesibilidad a los recursos genéticos microbianos de un ecosistema.
Durante el año 2021 se realizaron estudios en diferentes hábitats de la Serranía de la Lindosa (Guaviare), se tomaron muestras de agua (n=41) en caños y lagunas, suelos rizosféricos y no rizosféricos (n=60), en diferentes coberturas vegetales (bosques naturales primarios y secundarios, sabanas naturales, herbazales inundables y no inundables). Con estas muestras, se caracterizó la diversidad bacteriana por Illumina Miseq.
En los ecosistemas acuáticos, se observaron diferencias en la abundancia de organismos del dominio Bacteria entre caños y lagunas, siendo menor el número de ASVs (amplicon sequence variant) en estas últimas, sugiriendo menor diversidad. Se observó predominio, tanto en los caños con en las lagunas, de los Filos Proteobacteria, Actinobacteria y Cyanobacteria. Se profundizó en el análisis de abundancia y diversidad de Cianobacterias debido a que son un grupo diverso de procariotas fotosintéticos aerobios, que exhiben una fisiología versátil y amplia tolerancia ecológica para habitar con éxito en una amplia variedad de ambientes iluminados, tanto planctónicos como bentónicos (Cohen & Gurevitz, 2006; Sharma et al., 2014; Waterbury, 2006). Se analizaron 44 muestras de agua tomadas en las cuencas: Delicias – Laguna Negra (sectores: cascada Las Delicias, Puerto Arturo y Laguna Negra); Orión – María (sectores: Finca el Conuco, Laguna Chica y Laguna Grande). Se encontró que el 1.2% de la comunidad microbiana corresponde a Cianobacterias (589 ASVs), siendo mayoritarias en caños que en lagunas. Respecto a la resolución taxonómica, hay abundancia de las clases: Oxyphotobacteria, Melainabacteria y Sericytochromatia. A nivel de género predominaron Cyanobium PCC-6307 y Leptolyngya PCC-6306. El análisis de diversidad beta, indica que variables como el pH, acidez total, demanda biológica de oxígeno (DBO) y el potencial redox, influyen en la composición de la comunidad microbiana en caños y lagunas.
En los suelos se observó mayor diversidad alfa en suelos no rizosféricos que en suelos rizosféricos. A nivel taxonómico no se encuentran cambios significativos relacionados con la composición de las comunidades, detectando predominio de los Filum Proteobacteria, Acidobacteria y Actinobacteria. La diversidad beta permitió inferir que las variables fisicoquímicas asociadas a la saturación de magnesio y sodio, el pH y la conductividad eléctrica fueron las que separaron las muestras por sectores, lo que significa que las comunidades bacterianas del suelo se diferenciaron más por su posición geográfica o tipo de cobertura, que por su cercanía a sistemas radiculares específicos.
En el 2022 se realizó un muestreo el alto río Amazonas en la rivera del municipio de Leticia, complejo lagunar de Yahuarcaca. Se tomaron muestras de suelos aluviales (n=15) y se observó que la composición microbiana está conformada por representantes del Filum Proteobacteria, de las clases Alpha, Delta y Gamma, seguido por los Filum Acidobacteria, Actinobacteria, Cloroflexi y Verrucomicrobia.
Aislamiento e identificación de bacterias funcionales.
Durante los muestreos realizados entre los años 2019 y 2020 en diferentes puntos del departamento de Caquetá y de la Serranía de la Lindosa en el departamento de Guaviare, se han empleado diferentes estrategias para el aislamiento de microorganismos con diferentes características funcionales, con el fin de conservar ex situ el recurso genético microbiano y facilitar la ampliación del conocimiento sobre la biodiversidad microbiana y su uso en procesos de biorremediación, restauración ecológica y bioprospección. A partir de aguas, sedimentos y suelos, se han aislado, identificado y conservado microorganismos de los géneros Rhodococcus, Streptomyces, Gordonia, Nocardia, Agrococcus, Arthrobacter, Arsenicicoccus, Serratia, Acinetobacter, Brevundimonas, Enterococcus, Priestia, Bacillus, Paenibacillus, Niallia, Pseudomonas, Burkholderia, Shewanella, Chromobacterium, Microbacterium, Pantoea, Achromobacter, Mesorhizobium, Lysinibacillus, Leclercia, Stenotrophomonas y Sinomonas entre otras.
Para algunas de estas cepas, se realizó la secuenciación del genoma, por tecnología Illumina Hiseq y/o tecnología Nanopore, con el fin de elucidar su afiliación taxonómica. En la actualidad se cuenta con información genómica de las cepas Pseudomonas sp. COLMIS 53B, Agrococcus sp COLMIS 47B, Kitasatospora sp. COLMIS 38A, Burkholderia sp. COLMIS 51B, Chromobacterium sp. COLMIS 58B, Pseudarthrobacter sp COLMIS 348, Gordonia sp. COLMIS 303, Arthrobacter sp. COLMIS 329, Arsenicicoccus sp. COLMIS 466, Rhodococcus sp. COLMIS 296, Streptomyces sp. COLMIS 82B, Burkholderia sp. COLMIS 325, Pseudomonas sp. COLMIS 151, Pantoea sp. COLMIS 324, Bacillus sp. COLMIS 139, Acinetobacter sp. COLMIS 101-B, Microbacterium sp. COLMIS 121, Pseudomonas sp. COLMIS 142, Leclercia sp. COLMIS 114, Stenotrophomonas sp. COLMIS 115, Acinetobacter sp. COLMIS 119, Pseudomonas sp. COLMIS 147, Bacillus sp. COLMIS 172, Ralstonia sp. COLMIS 200 y Burkholderia sp. COLMIS 204.
A partir de los datos genómicos se ha realizado el análisis de los índices generales de relación genómica (OGRI) que incluyen la identidad promedio de nucleótidos y la hibridación digital de ADN-ADN (dDDH), esto ha permitido sugerir que las cepas COLMIS 47B, COLMIS 114 y COLMIS 329 pueden ser nuevas especies de los géneros Agrococcus, Leclercia y Arthrobacter respectivamente.
Estas cepas ingresaron a la colección de microorganismos del Instituto SINCHI – COLMIS (RNC No. 282). A la fecha, COLMIS cuenta con un total de 466 depósitos, de los cuales 16 son cepas de referencia. La colección cuenta con 437 cepas bacterianas y 29 levaduras. Para el caso de bacterias, predominan los géneros Bacillus y Pseudomonas, y para las levaduras predominan los géneros Wickerhamomyces e Issatchenkia. Respecto a los grupos funcionales, predominan las bacterias resistentes a Hg, las bacterias solubilizadoras de fosforo y fijadoras de nitrógeno de forma libre (diazótrofas). Actualmente, se realiza la preservación de los microorganismos según lineamientos de calidad establecidos por la WFCC (World Federation for Culture Collection). COLMIS cuenta con una aplicación Web en lenguaje de programación PHP/Python/JavaScript para administrar la hoja de vida de los registros de la colección. Este desarrollo se desplegó en el servidor del laboratorio de Biotecnología y Recursos Genéticos del I. Sinchi y permite el ingreso, la consulta y la actualización de la información consolidada. El sistema se desplegó en la siguiente url: http://colmis.sinchi.org.co/
Finalmente, y con el fin de contribuir a los inventarios de la biodiversidad, se realizó el depósito de 71 cepas de bacterias aisladas de los departamentos de Caquetá, Amazonas y Vaupés en el SIB Colombia (red nacional de datos abiertos sobre biodiversidad) y de GBIF (infraestructura mundial de información en biodiversidad) a través del DOI: https://doi.org/10.15472/qfnt6t.
Recomendaciones recursos genéticos y microorganismos
- Hacer investigación sobre el aporte de los microorganismos en los ciclos biogeoquímicos y su relación con el cambio climático.
- Incluir estudios funcionales de las comunidades microbianas haciendo uso de herramientas moleculares como la metagenómica y transcriptómica.
- Continuar con la secuenciación de genomas de microorganismos aislados a partir de diferentes muestras ambientales.
Recursos Genéticos de Flora y Fauna
Las secuencias barcode de especies de flora, ha sido generada a partir de 1622 muestras (aproximadamente), de tejido vegetal colectadas en diferentes sitios de muestreo de la región Amazónica. A partir de estas muestras se ha generado hasta el año 2019, un total de 2730 secuencias depuradas de marcadores barcode (Matk y Rcbl), ITS, microsatélites, entre otros. La mayoría de las secuencias (39% y 52%), corresponden a los marcadores de DNA plastidico Matk y Rcbl respectivamente. En términos de abundancia de secuencias por grupo taxonómico, el total de secuencias depuradas comprende 89 familias, 247 géneros y 451 especies. Las familias de plantas que cuentan con un mayor número de secuencias son: Cyatheaceae 322 (14%), Burceraceae 231 (10%), Euphorbiaceae 194 (8%), Bromeliaceae 145 (6%). Los géneros de especies vegetales con mayor representación de secuencias generadas son: Cyathea 275 (12%) y Potium 120 (5%). En el caso de especies vegetales, la mayoría de las secuencias 336 (14%), no corresponden a ningún grupo taxonómico. Así mismo, las especies con mayor número de secuencias con Cariniana pyriformis 84 (4%) y Cedrela odorata 53 (2%). Lo cual sugiere la falta de poder de resolución en la identificación de especies a partir de las secuencias de los marcadores moleculares evaluados.
Por otro lado, y con el fin de resolver los problemas asociados a la resolución taxonómica de especies maderables, basada en el uso de códigos de barras de ADN, se optó por utilizar la secuencia completa del cloroplasto como código de barras (“Super-barcode”) (Wicke and Schneeweiss, 2015). La secuencia completa del genoma de cloroplasto- cp en plantas, tiene una longitud de 110 a 160 kb, la cual excede significativamente cualquier secuencia barcode y provee una mayor variación para discriminar especies cercanas con morfortipos parecidos en estudios filogenéticos, filogeográficos y poblacionales (Straub et al., 2012). Así mismo, el genoma cp tiene una alta tasa de variación interespecífica (i. e. alto número de copias), y baja variación intraespecífica (i.e. baja sustitución nucleotidica y recombinación), por lo cual es el marcador barcode genómico más apropiado (Twyford y Ness, 2017; Sheunert et al., 2020). En el año 2020, se priorizo la generación y estandarización de un protocolo de extracción de DNA de alto peso molecular (HMW), en la especie C. odorata (especie maderable y la secuenciación por Nanopore en el equipo minION de individuos de C. odorata representativos, de las diferentes poblaciones colombianas, con el fin de generar genomas cp de referencia que puedan ser utilizados como superbarcodes o para generar nuevos marcadores moleculares específicos. Se obtuvo DNA-HMW, utilizando el protocolo reportado por Schalamun y Schwessinger (2017), para especies del género Eucalyptus y adaptado en el Laboratorio de Recursos Genéticos de SINCHI para C. odorata, a partir tejido foliar disponible en el COAH. En el 2021 se realizó la secuenciación por la tecnología nanopore en el equipo minION del I. Sinchi del genoma cp de dos individuos de C. odorata (OGP2096 y OGP2146). A partir de estas secuencias, se identificaron cuatro (4) SNPs tipo deleción, los cuales pueden ser incluidos en estudios de genotipificación de poblaciones de C. odorata en Colombia e.g., mediante el uso de la técnica de PCR convencional, qPCR (PCR cuantitativa en tiempo real y/o ddPCR (PCR digital) en el Instituto SINCHI, con el fin de generar una herramienta de huella genómica propia de estas poblaciones. Para el 2022 se estandarizó el protocolo de amplificación por PCR convencional de 4 variantes estructurales (SNPs) en muestras de ADN de Cedrela odorata y se identificaron otros 5 SNPs candidatos por evaluar en estudios de genética poblacional de cedro. Así mismo, se optimizó el proceso de extracción de ADN a partir de 98 muestras de tejido foliar de C. odorata, para continuar el proceso de secuenciación MASSArray con el fin de identificar una mayor cantidad de SNPs que permitan determinar la estructura poblacional de cedro en el país.
En el 2022, en el área de Flora se priorizaron para la obtención de secuencias barcode, 16 especies de plantas conservadas en el herbario amazónico colombiano (COAH), caracterizadas por presentar alta presión de tráfico o con requerimientos de identificación para levantamientos de vedas (Maderables y Bromelias). Se amplificaron y secuenciaron 28 códigos de barra de cloroplasto (rbcL y matK) con los cuales se confirmó la correcta asignación taxonómica a nivel de genero estos individuos. Posteriormente con el fin de aportar al inventario nacional de los recursos genéticos de flora amazónica del país, se asociaron 323 secuencias de códigos de barra de flora alojados en BOLDSystems, a la red nacional de datos abiertos sobre la biodiversidad (SiB). En el área de Fauna, se estandarizó la reacción de amplificación por PCR convencional de los marcadores moleculares 16S, COI y Cyt B en 38 muestras de tejido de anfibios y mamíferos. Como resultado se obtuvieron 132 secuencias de códigos de barra de ADN mitocondrial, las cuales, al compararlas con las bases de datos disponibles (NCBI) coinciden en su mayoría con la asignación taxonómica a nivel de género. Sin embargo, al presentar una alta variabilidad y poca similitud a nivel de especie, las secuencias obtenidas se implementarán en análisis filogenéticos con el fin de sustentar la asignación de nuevas especies de anfibios de los géneros Bolitoglossa y Boana, y de murciélagos, del género Anoura de la Amazonia colombiana.
Finalmente, se dio inicio al desarrollo de estudios de Metabarcoding en plantas, los cuales permiten la identificación de múltiples especies a partir de muestras ambientales de origen mixto. Para esto, se desarrolló un protocolo de extracción de ADN a partir de muestras de polen, colectado de aves y murciélagos. La eficiencia del protocolo en la obtención de ADN de alta calidad permitirá la obtención de secuencias de ADN correspondientes a códigos de barra, por medio de tecnologías de secuenciación de última generación (Nanopore-MinION)
Recomendaciones Recursos Genéticos de Flora y Fauna
De acuerdo con las necesidades taxonómicas y sistemáticas de las especies de fauna disponibles en las colecciones biológicas, se priorizarán anualmente taxones para avanzar en información genética que permita refinar la curaduría, definir límites de distribución de los distintos linajes para los análisis de diversidad y estrategias de conservación en la Amazonia.
Recomendación para flora????
Biología del suelo
Organismos del suelo
Uno de los recursos de investigación en el instituto Sinchi ha sido el estudio de la biología de los suelos amazónicos. En general se acepta que el 70% de los suelos amazónicos son ácidos y de baja fertilidad (Quesada et al. 2010), lo cual hace que las diferentes comunidades biológicas del suelo juegen un papel crucial en la descomposición de la materia orgánica y por ende en la nutrición de los ecosistemas amazónicos. Esta investigación se ha basado en el conocimiento de dos grupos biológicos: Los hongos formadores de micorrizas arbusculares y la macrofauna del suelo.
Los hongos son un grupo diverso en los suelos que gobiernan el ciclaje de carbono y la nutrición vegetal. En estudios globales se ha estimado que aproximadamente el 85.4% de los OTUs (unidades taxonómicas operativas) obtenidos en muestras de suelo corresponden a hongos (Tedersoo et al. 2014). Se dice que la relación planta-hongo en la naturaleza puede ser de 1:8±10 (Hawksworth 2001), pero la diversidad de hongos asociados a las plantas y por ende la proporción planta-hongo varía geográficamente. En la región tropical y especialmente en la Amazonía los hongos formadores de micorrizas tienen un papel fundamental en la nutrición vegetal. El principal beneficio, para la planta, que le confiere la micorriza arbuscular es una mayor asimilación de fósforo (P), elemento importante en la síntesis proteica y en la producción de energía (ATP). Adicionalmente, la micorriza arbuscular también mejora la absorción de microelementos como el zinc (Jansa et al. 2003; Marschner y Dell 1994), le confiere mayor resistencia contra patógenos a la planta (Newsham et al. 1995; Vaast et al. 1998), mayor tolerancia a la sequía (Davies et al. 1992) y mayor tolerancia a la toxicidad de aluminio y magnesio en el suelo (Cuenca 2001). Además, los hongos formadores de micorrizas arbusculares (HMA) trabajan sinérgicamente con bacterias fijadoras de nitrógeno y promotoras del crecimiento, generando un efecto conjunto positivo que incide directamente en una mejor nutrición de las plantas.
Adicionalmente a la función que cumplen las micorrizas arbusculares en la nutrición de las plantas, los HMA también tienen un papel importante en el mantenimiento de la estructura del suelo y la prevención contra la erosión, así como en la detoxificación de los suelos. Estos hongos producen una proteína denominada glomalina, la cual actúa como un pegante (Miller y Jastrow 2000), adhiriendo materia orgánica y formando agregados biológicos estables en el suelo. También, el mismo micelio del hongo actúa como un microtaladro en el suelo creando canales y aumentando la microporosidad del suelo. Se ha observado que los HMA son capaces de colonizar plantas en ambientes contaminados por arsénico (Bona et al. 2011), antraceno (Debiane et al. 2008) y fenol (Ibañez et al. 2011), teniendo así un papel importante en la biorremediación de suelos expuestos a agroquímicos, a contaminantes industriales y a suelos degradados por minería. Por tal motivo, se considera que los hongos formadores de micorrizas arbusculares tienen un papel más preponderante en los suelos que otros grupos de hongos y que su rol es multifuncional.
Por su parte, la macrofauna edáfica, 111compuesta principalmente por artrópodos terrestres que llevan todo su ciclo de vida en el suelo o que sólo están en el suelo en algún estaiío de su vida. La macrofauna edáfica puede ser clasificada según su rol en el ecosistema, grupos alimenticios o funcionales y su tamaño. La macrofauna del suelo es en su mayoría detritívora. En este grupo están las termitas (Isoptera), las hormigas (Hymenoptera), los escarabajos y cucarrones (Coleoptera), las cochinillas de tierra (Isopoda), los milpiés (Diplopoda) y algunas larvas de insectos principalmente Coleoptera y Lepidoptera. El otro grupo importante de la macrofauna edáfica lo constituyen los depredadores, donde se incluyen los ciempiés y arácnidos grandes (Moreira et al. 2012; Swift et al. 1979; Swift et al. 2012). Las hormigas y las termitas son capaces de modificar la estructura del suelo al crear galerías, nidos, cámaras y bolas de excremento que cambian la aireación, la retención de agua, la infiltración y el drenaje (De la Rosa y Negrete-Yankelevich 2012), por ello, junto con las lombrices de tierra, estos tres grupos animales son considerados los “ingenieros del suelo”. Se estima que más del 80 % de los artrópodos terrestres que habitan los suelos tropicales no han sido descritos por la ciencia, razón por la que tampoco se conocen detalles de su ecología (Wall et al. 2010).
Resultados
1) Avances en el conocimiento de los hongos formadores de micorrizas arbusculares
Actualmente, se cuenta con una base de datos que incluye 1.268 registros, provenientes de 855 muestras de suelos a las cuales se ha realizado la determinación morfológica a partir de las esporas de los hongos encontrados, y 413 muestras procesadas molecularmente y de las cuales se han obtenido secuencias específicas de estos hongos para su determinación. De estas, el 70% corresponde a información proveniente de muestras de suelo y el 30% proveniente de muestras de raíces de plantas. A partir de esta información, el Instituto Sinchi es la entidad colombiana con mayor información sobre hongos formadores de micorrizas arbusculares (HMA) del país, en ambientes naturales.
Los hongos formadores de micorrizas forman asociaciones con las plantas que juegan un papel importante en su nutrición y en el mantenimiento de suelos saludables. En la Amazonía, dados los bajos niveles de fósforo que presentan buena parte de sus suelos, esta asociación adquiere una mayor relevancia en la nutrición de las plantas. En el caso específico de la Amazonía colombiana se han reportado 156 taxones virtuales (unidades taxonómicas filogenéticamente definidas) de hongos formadores de micorrizas arbusculares (HMA) que corresponden al 49 % de la diversidad mundial y al 68 % de la diversidad reportada para los trópicos (Peña-Venegas 2021).
Se ha observado que, en la Amazonía colombiana, existe una mayor abundancia de esporas de HMA en bosques secundarios y pasturas que en bosques primarios (Peña-Venegas et al. 2006), por cuanto los recuentos y estimación de la diversidad por métodos que dependen de la recuperación de esporas del suelo puede generar interpretaciones equivocadas sobre la diversidad y abundancia de géneros y especies que esporulan poco o que no tienen el mecanismo de esporular como una respuesta ante condiciones de estrés en el suelo.
La comunidad de HMA en suelos amazónicos se caracteriza por la dominancia de las familias Glomeraceae, Acaulosporaceae y Gigasporaceae (Freitas et al. 2014). Los géneros más diversos y representativos en los suelos de la región amazónica son Glomus y Acaulospora (Martin et al. 2001; Caproni et al. 2003; Peña-Venegas et al. 2007; Stürmer y Siqueira 2011), una condición que parece repetirse a lo largo de todos los trópicos (Marinho et al. 2018). Es interesante observar que, independientemente de la metodología usada para determinar la composición de las comunidades de estos hongos, la proporción de los géneros no cambia. El género Glomus es el género dominante en los suelos amazónicos.
La composición de la comunidad de estos hongos no es homogénea a lo largo de la región amazónica, y aún cuando se ve influenciada por la composición de los suelos y la flora, su composición no está directamente relacionada con estos, por cuanto no existe un patrón composición florística-micorrizas o composición suelo-micorrizas (Peña-Venegas et al. 2021).
Las plantas no son colonizadas por una sola especie de HMA. En general, son micorrizadas por un conjunto de especies de HMA que varía entre especies de plantas, ecosistemas y usos del suelo. Se ha reportado que las plantas en los trópicos albergan un mayor número de HMA que las de ecosistemas templados (Öpik et al. 2006). Especies de plantas de un bosque templado pueden albergar entre 8 y 15 especies (como VT) en sus raíces (Saks et al. 2014) con un promedio de 5,6 VT por planta (Öpik et al. 2006), mientras plantas de bosques tropicales albergan en sus raíces un promedio entre 18,2 -19 VT por planta (Öpik et al. 2006; Peña-Venegas et al. 2014).
El uso de técnicas moleculares para el estudio de estos organismos permitió evidenciar que, existe un cambio en la composición de las comunidades de hongos formadores de micorrizas arbusculares con el tipo de cobertura: Coberturas de gramineas introducidad (pastos) son dominadas por Glomus, pero durante su regeneración natural y paso a convertirse de suevo en coberturas boscosas, la comunidad de hongos deja de ser dominada por Glomus y para a ser dominada por Paraglomus, siendo éste un género asociado dominantemente a coberturas amazónicas boscosas (Peña-Venegas et al. 2021).
Los resultados obtenidos por el Instituto Sinchi en los últimos años, hizo que en el año 2019, la red de investigadores en micorrizas de sur América, postulara al Instituto Sinchi como la entidad anfitriona.
Recomendaciones
Ya se ha venido trabajando en la estimación de la biodiversidad de hongos formadores de micorrizas arbusculares en el Amazonas a lo largo de mas de 20 años. Sin embargo, tras nuevos muestreos el número de nuevos géneros, especies y taxones virtuales sigue siendo reportado. Esto lo que indica es que aún estamos lejos de tener los inventarios completos de estos hongos para la Amazonia colombiana.
Los HMA han sido reportados como especies de importancia en la restauración de ecosistemas contaminados o disturbados y en el mantenimiento del stock de carbono en el suelo. Se han comenzado a dar algunos pasos en la dirección de entender el rol que estos hongos cumplen en ecosistemas amazónicos afectados por minería, otros contaminantes o la deforestación, y cómo responden ante el cambio climático. Sin embargo, es necesario poder avanzar en cómo aplicar el conocimiento de estos hongos a la resolución de estos problemas y la conservación de los ecosistemas.
2) Avances en el conocimiento de la macrofauna del suelo
En los últimos años se ha evidenciado el importante rol que desempeña la macrofauna en la estructura del suelo (Decaëns et al. 1999, Barros et al. 2001, Mathieu et al. 2005, Bignell et al. 2012, Velásquez et al. 2012), en especial con las lombrices, termitas y hormigas, llamados “ingenieros del ecosistema” (Lavelle 1997, Decaëns et al. 1999). Estos grupos ayudan a mantener la productividad del suelo debido a que intervienen directamente en los procesos de reciclaje y descomposición de nutrientes. Igualmente se ha evidenciado que estos grupos modifican las propiedades físicas del suelo (Mathieu et al. 2005). Así, la generación de agregados y estructura del suelo cambia dependiendo de si las condiciones son favorables o no para la macrofauna (Lavelle y Spain 2001).
El instituto Sinchi cuenta con una base de datos con registros de 13.486 especímenes de 12 clases, 53 órdenes, 246 familias, 231 géneros y con alrededor de 1.600 morfoespecies descritas. Estos registros indican que la Amazonia colombiana es la región más biodiversa en hormigas y termitas del pais, de acuerdo a los inventarios registrados y publicados por el Instituto Sinchi.
De los estudios realizados en la Amazonía se ha encontrado que las hormigas, los colémbolos y los coleópteros son los grupos más abundantes, exhibiendo una heterogeneidad de hábitos alimenticios y nichos, con predominancia de los detritívoros, los cuales actúan principalmente transformando la hojarasca (Cárdenas et al. 2017).
Se sabe la riqueza, la abundancia y la diversidad de estos organismos se ven altamente influenciadas por los diferentes cambios de uso de suelo (Suárez et al. 2015). Por ello, la macrofauna edáfica se ha propuesto como indicadora de estados de perturbación dada su respuesta a las diversas alteraciones antrópicas al medio ambiente (Barros et al. 2001).
Para el estudio de la macrofauna de los suelos amazónicos, se ha seguido la metodologìa TSBF (anderson & Ingram 1993), la cual es una técnica validada mundialmente y que se basa en la colecta de todos los organismos visibles en monolitos de suelo de 25 x 25 x 30 cm. A partir de estos estudios se ha podido establecer que las comunidades de macrofauna edáfica están compuestas por grupos taxonómicos clasificados entre 15 y 22 grupos con gran contraste ecológico (Barros et al. 2008). Entre las clases más importantes encontramos Formicidae, Coleoptera, Isoptera, larvas, Chilopoda, Diplopoda, Blattodea, Hemiptera, Diplura, Isopoda, Opiliones, Orthoptera, Pseudoscorpionida, Schizomida y Dermaptera, además de otros organismos no tan diversos en suelos como Mantodea y Phasmatodea, y otros que no son fáciles de recolectar en monolitos de suelo como Onichophora, Scorpiones y Ricinulei.
Aproximadamente por cada monolito de 25 x 25 x 30 cm se pueden encontrar doce morfoespecies. Las hormigas son el grupo más rico con cuatro especies en promedio por monolito. Los coleópteros y las termitas tienen en promedio una especie. Los demás grupos tienen una media inferior a una morfoespecie por monolito. Estos valores obtenidos para suelos de la Amazonia colombiana son un poco más bajos que los reportados para suelos de bosques amazónicos brasileños (Barros et al. 2004, 2008). Los suelos amazónicos pueden llegar a albergar en total una densidad media de 495 individuos de macrofauna edáfica por metro cuadrado. Los ecosistemas naturales presentan la mayor densidad con una media de 626 individuos en bosques de vegetación secundaria y 557 individuos en bosques maduros; por otra parte, en las coberturas de suelos perturbados y degradados la densidad puede llegar a disminuir desde un 51 % hasta un 12 %, donde las pasturas comprenden una media de 490 y los rastrojos 322 individuos.
Otra variable que viene siendo evaluada es la densidad y su relación con la bimasa del suelo. La densidad más alta corresponde al bosque secundario, que por su proceso de sucesión con recambio de especies y buena disponibilidad de alimento es propicio para que diversos organismos puedan existir. Esto puede traducir en grupos muy densos que aportan un numero significante de biomasa al suelo. Sin embargo, hay grupos que son muy pocos densos e igual pueden estar aportando una cantidad significativa de biomasa como algunos grupos de arachnida y miriapoda (Barros et al. 2002, 2004).
De acuerdo a el tipo de cobertura muestreada y su estado de intervención, se ha podido establecer que la densidad de hormigas se mantiene constante, no siendo afectadas por cambios en las cobereturas (Rodriguez et al. 2021). Otros grupos son mucho más sensibles al cambio de cobertura, como es el caso de los coleópteros que se reducen en más de un 38 % en suelos perturbados; así como en las larvas de insectos y milpiés.
Colombia alberga actualmente un 9 % de las especies de hormigas del mundo, con 11 subfamilias registradas y 1200 especies reportadas (Antwiki 2016, Fernández et al. 2018). Del total de las 11 subfamilias presentes en Colombia, todas estuvieron presentes en la porción de la región Amazónica colombiana muestreada, lo cual es una alta representación para una sola región, comparada con el resto del territorio nacional.
Los suelos amazónicos albergan una gran cantidad de especies de termitas. La falta de estudios relacionados con estos organismos en el país ha hecho pasar desapercibida su gran biodiversidad. Los nuevos registros y nuevas especies que se han podido datar en los últimos años, abre las puertas a todo un nicho de trabajo para el estudio de las termitas, no sólo en la región amazónica sino en el país (Castro y Peña-Venegas 2018, Castro et al. 2018, Pinzon y Castro 2018). La composición de las comunidades de termitas en el suelo varia de las encontradas en los estratos epigeos y arbóreos donde otras especies de termitas nidifican (Dibog et al. 1999, Almeida et al. 2017). En el suelo se encuentran mayormente especies de la subfamilia Apicotermitinae que se caracterizan en el neotrópico por la ausencia de la casta del soldado y su identificación ha sido un problema taxonómico (Bourguignon 2016). En los últimos años el estudio de estas termitas ha podido esclarecer un poco más su taxonomía, en donde la descripción de nuevos géneros y las redescripciones de especies han permitido hacer una mejor identificación de estas especies (Bourguignon et al. 2010, 2016, Scheffrahn 2013, Acioli y Constantino 2015, Scheffrahn et al. 2017, Castro et al. 2018).
Estos resultados han permitido que en 2021 el Instituto Sinchi fuera invitado a participar en 4 proyectos internacionales colaborativos con los investigadores más reconocidos en macrofauna edáfica: SOILBON, SOILBON-GBIF, SOILFAUNA y SOILSERVICES.
Recomendaciones
Actualmente se cuenta con información taxonómica importante de hormigas y termitas de suelos amazónicos que antes no existía ni para la región ni para Colombia. Aún cuando se ha logrado un avance significativo en el estudio de hormigas y termitas, falta ampliar las investigaciones a coleópteros, arañas, isópodos y miriápodos, encontrando respuestas adicionales a cómo estos organismos están influenciando la composición y la estructura del suelo, además de su susceptibilidad al cambio de uso del suelo e intervenciones antrópicas.
Por otra parte, se viene avanzando en la determinación de especies indicadoras de disturbio del suelo, en cuantificar el aporte que estos organismos hacen al stock de carbono en el suelo y su relación con los servicios ecosistémicos del suelo y el cambio climático. Así, será posible aportar nueva información para la conservación de los suelos y los ecosistemas amazónicos.
Fichas de los resultado investigación científica en el marco del PICIA 2019-2023

Fichas de los resultado investigación científica en el marco del PICIA 2019-2022

1. Conocimiento para el uso, manejo y conservación de la diversidad biológica

2. Conocimiento tradicional y diálogo de saberes

3. 2022 Programa Sostenibilidad e Intervención

4. Estrategias de remediación y monitoreo de la contaminación de ambientes final

5. Desarrollo tecnológico e innovación – Biotecnología – Bioprospección
5. Desarrollo tecnológico e innovación – Biotecnología – Bioprospección (2019-2022)
6. Dinámicas sociambientales en la Amazonia

8. Modelamiento ambiental y escenarios dinámicos

9. Monitoreo y ordenamiento ambiental

10. Avances en restauración y propuesta de insumos para próximo PICIA

11. Paisajes Productivos Amazonia
