
Equipo: Clara P. Peña Venegas, Daniel Castro
El suelo es uno de los hábitats más diversos y complejos del planeta, con más del 40% de los organismos terrestres asociados a él en alguna etapa de su ciclo de vida (Jiménez et al., 2006; FAO et al., 2020; Orgiazzi et al., 2016). A pesar de su relevancia, la biodiversidad del suelo continúa siendo una de las menos conocidas, especialmente en regiones como la Amazonía colombiana. Esta región alberga ecosistemas únicos de alta complejidad ecológica, donde la biota del suelo desempeña un papel esencial en el sostenimiento de procesos clave como el reciclaje de nutrientes, la descomposición de la materia orgánica, el almacenamiento de carbono y la regulación hídrica.
Entre los principales grupos biológicos del suelo, la macrofauna edáfica representa un reservorio clave de biodiversidad. Compuesta por lombrices, insectos sociales (como termitas y hormigas) y otros invertebrados, esta fauna influye directamente en la estructura y funcionamiento del suelo tropical (Bottinelli et al., 2015; Castro et al., 2025). Estos organismos no solo fragmentan y redistribuyen la necromasa, sino que también modifican físicamente los perfiles del suelo y afectan las dinámicas ecológicas a distintas escalas (Griffiths et al., 2019; Andriuzzi & Wall 2018). A pesar de su importancia, la macrofauna es frecuentemente subestimada en los estudios de biodiversidad (Gongalsky 2021).
Las micorrizas arbusculares (AM) desempeñan un papel clave en los suelos tropicales, ya que mejoran la capacidad de las plantas para absorber nutrientes, en especial fósforo. Asimismo, fortalecen la resistencia al estrés hídrico y favorecen la formación y estabilidad de agregados en el suelo (Li et al., 2020; Peña-Venegas & Cardona, 2010). En la región amazónica, estas asociaciones simbióticas han demostrado ser cruciales para procesos como la regeneración natural de los bosques, la conservación de la diversidad vegetal y la capacidad del ecosistema para recuperarse frente a perturbaciones (Peña-Venegas et al., 2022). Su diversidad y abundancia se ven condicionadas por variables como la cobertura vegetal, las características texturales del suelo y el tipo de manejo del territorio, lo que las posiciona como posibles bioindicadores del estado ecológico del suelo en la Amazonia (Brussaard et al., 2007).
Los suelos de la Amazonía colombiana, predominantemente Oxisoles y Ultisoles de baja fertilidad (IGAC 2012), dependen en gran medida de la actividad biológica para su sostenibilidad. Sin embargo, estos suelos enfrentan crecientes presiones por actividades antrópicas como la deforestación y el cambio de uso del suelo, que generan focos de degradación acelerada. La escasa información a escalas semidetalladas y detalladas (1:25.000) sobre sus propiedades, biodiversidad y funciones limita la toma de decisiones informada para su conservación y uso sostenible (Minambiente 2016).
Entre los principales grupos biológicos del suelo, la macrofauna edáfica representa un reservorio fundamental de biodiversidad. Está compuesta por invertebrados como lombrices, coleópteros, miriápodos y, especialmente, insectos sociales como hormigas y termitas, que constituyen una porción significativa de la biomasa del suelo en los ecosistemas tropicales (Barros et al., 2002; Fittkau & Klinge 1973). Estos organismos tienen un rol protagónico en la formación y estabilización de la estructura del suelo, así como en el fraccionamiento y movilización de la necromasa (Andriuzzi & Wall 2018; Griffiths et al., 2019). Las hormigas, por su diversidad funcional y patrones de dominancia ecológica, y las termitas, por su capacidad de digestión de celulosa y construcción de galerías, son ingenieros del ecosistema que afectan múltiples funciones del suelo (Decaëns et al., 1994; Gerlach et al., 2013; Forstall-Sosa et al., 2021). No obstante, la macrofauna sigue siendo uno de los componentes más subestimados de los ecosistemas terrestres (Gongalsky 2021).
Frente a este escenario, el Instituto Amazónico de Investigaciones Científicas – SINCHI ha priorizado el estudio de la biología del suelo como parte de su estrategia para fortalecer el conocimiento sobre la biodiversidad amazónica. Particularmente, se han desarrollado estudios sobre hongos micorrízicos arbusculares, fundamentales para la nutrición vegetal en suelos empobrecidos, y sobre la macrofauna del suelo, destacando el uso de metodologías estandarizadas como el protocolo TSBF (Anderson & Ingram 1993) para el muestreo y análisis de comunidades edáficas. Estos estudios integran técnicas morfológicas y moleculares que permiten avanzar en la caracterización taxonómica y funcional de estos organismos, aportando evidencia clave para la planificación territorial, el manejo sostenible del suelo y la formulación de políticas públicas de conservación.
Estado del conocimiento de los recursos naturales del suelo
Hongos formadores de micorrizas arbusculares
En el primer semestre de 2025 se realizó la actualización de la base de datos, la cual cuenta con 1104 muestras que fueron analizadas para determinar la presencia de géneros/especies de estos hongos por técnicas tradicionales a partir de las morfologías de las esporas, y con 436 muestras que fueron analizadas para determinar la presencia de géneros/especies de estos hongos por técnicas moleculares. De estos análisis se tiene registro de 397 morfoespecies y taxones virtuales diferentes para la Amazonia colombiana. Se han podido identificar 21 géneros en la Amazonia colombiana: Claroideoglomus (con un morfotipo y 4 taxones virtuales), Glomus (con 120 morfotipos y 141 taxones virtuales), Funneliformis (con 8 morfotipos), Septoglomus (con 4 morfotipos y 1 taxón vitual), Simiglomus (con un morfotipo), Siverdingia (con un morfotipo), Sclerocarpum (con un morfotipo), Rhizophagus (con 4 morfotipos y 3 taxones virtuales), Acaulospora (con 31 morfotipos y 15 taxones virtuales), Entrophospora (con un morfotipo), Kuklospora (con dos morfotipos y un taxón virtual), Otospora (con un morfotipo), Cetraspora (con dos morfotipos), Diversispora (con un morfotipo y 4 taxones virtuales), Dentiscutata (con dos morfotipos), Gigaspora (con 4 morfotipos y un taxón virtual), Racocetra (con 2 morfotipos), Scutellospora (con 11 morfotipos y 5 taxones virtuales), Ambispora (con 2 morfotipos y 2 taxones virtuales), Paraglomus (con 14 taxones virtuales) y Archaeospora (con 11 taxones virtuales). Los dos últimos géneros solo han sido determinados por métodos moleculares. La abundancia de familias, géneros y especies se han reportado en los diferentes departamentos tal como se resume en la tabla 1. Estos resultados confirman que la determinación de los inventarios de estos hongos es necesario realizarlos usando las dos metodologías existentes ya que cada una se aproxima de manera diferente a la población existente en el lugar.
| Datos registros hongos formadores de micorrizas arbusculares | Semestre I 2025 |
Acumulado |
| TOTAL NÚMERO DE REGISTROS AMAZONIA | 10 | 397 |
| TOTAL NÚMERO DE ESPECIES AMAZONIA | 1 | 58 |
| # Familia AMAZONAS | 5 | 5 |
| # Familia CAQUETÁ | 0 | 6 |
| # Familias GUAINÍA | 0 | 6 |
| # Familias GUAVIARE | 0 | 6 |
| # Familias VAUPÉS | 0 | 4 |
| # Familias PUTUMAYO | 0 | 3 |
| # Familias sur META | 0 | 0 |
| # Familias sur VICHADA | 0 | 4 |
| # Familias BOTA CAUCANA | 0 | 0 |
| # Géneros AMAZONAS | 5 | 18 |
| # Géneros CAQUETÁ | 0 | 18 |
| # Géneros GUAINÍA | 0 | 14 |
| # Géneros GUAVIARE | 0 | 9 |
| # Géneros VAUPÉS | 0 | 12 |
| # Géneros PUTUMAYO | 0 | 9 |
| # Géneros sur META | 0 | 0 |
| # Géneros sur VICHADA | 0 | 5 |
| # Géneros BOTA CAUCANA | 0 | 0 |
| # especies AMAZONAS | 10 | 37 |
| # especies CAQUETÁ | 0 | 52 |
| # especies GUAINÍA | 0 | 24 |
| # especies GUAVIARE | 0 | 13 |
| # especies VAUPÉS | 0 | 23 |
| # especies PUTUMAYO | 0 | 13 |
| # especies sur META | 0 | 0 |
| # especies sur VICHADA | 0 | 5 |
| # especies BOTA CAUCANA | 0 | 0 |
Tabla 1. Resumen de los reportes de hongos formadores de micorrizas arbusculares por departamento.
Se destaca la identificación de 10 nuevos taxones virtuales de los generos Paraglomus (SS-PG1), Glomus (VT429, VT331, VT120, VT256, VT329, VT88 y VT291), Archaeospora (SS-Ar1 y VT338), y la especie Dentiscutata scutata descrita a partir de esporas aisladas de suelos.
La composición micorrícica de cada tipo de suelo es característica, teniendo especies de HMA que solo han sido reportadas en un ambiente edáfico determinado, pero que en su gran mayoría son compartidas por más de un tipo de suelo (Figura 1).
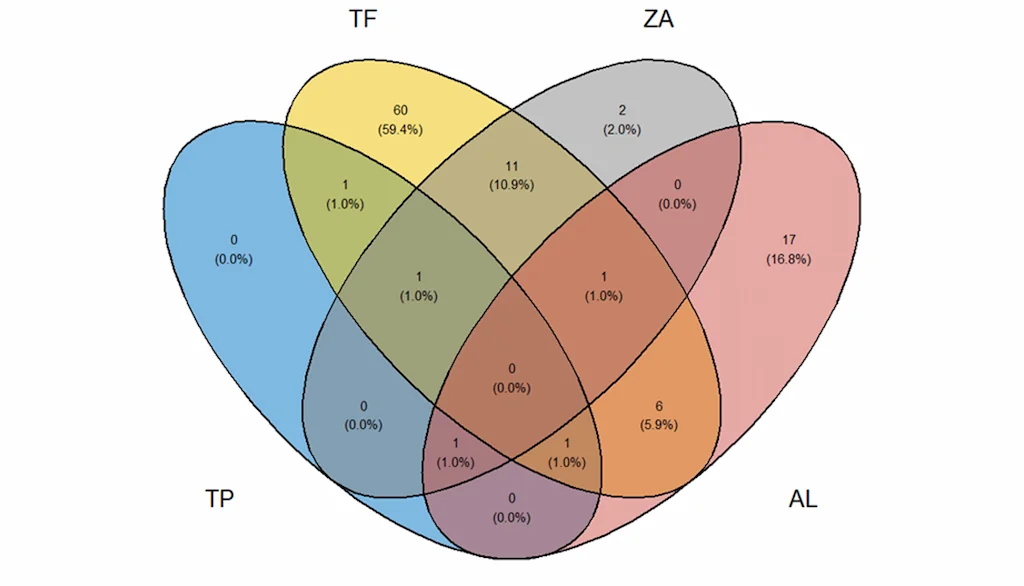
Figura 1. Diagrama de Venn indicando las morfo-especies de hongos formadores de micorrizas arbusculares en cada tipo de suelo: TF (suelos de tierra firme), ZA (suelos de arenas blancas); TP (suelos antropogénicos); AL (suelos aluviales). Fuente: Programa Ecosistemas y Recursos Naturales- Instituto SINCHI 2025.
A pesar de la importancia que los hongos formadores de micorrizas arbusculares tienen en el mantenimiento de los ecosistemas y que se resumen en las contribuciones al objetivo de Desarrollo sostenible de “Vida en la tierra” del artículo publicado por Stewart et al. (2025), se muestra que existe un poco reconocimiento de la importancia que estos hongos tienen en la preservación de los ecosistemas.
Macrofauna del suelo
Entre 2016 y 2024, el Instituto Amazónico de Investigaciones Científicas – SINCHI ha llevado a cabo un extenso inventario de la macrofauna del suelo como parte de su estrategia para caracterizar la biodiversidad edáfica en la Amazonia colombiana. Durante este periodo, se han colectado 675 monolitos TSBF, distribuidos en 135 sitios de muestreo en cinco de los seis departamentos de la región (Figura 2). Este esfuerzo ha generado más de 8.700 muestras edáficas (aproximadamente 2.600 por profundidad), permitiendo una caracterización detallada de la composición taxonómica y funcional de la macrofauna.
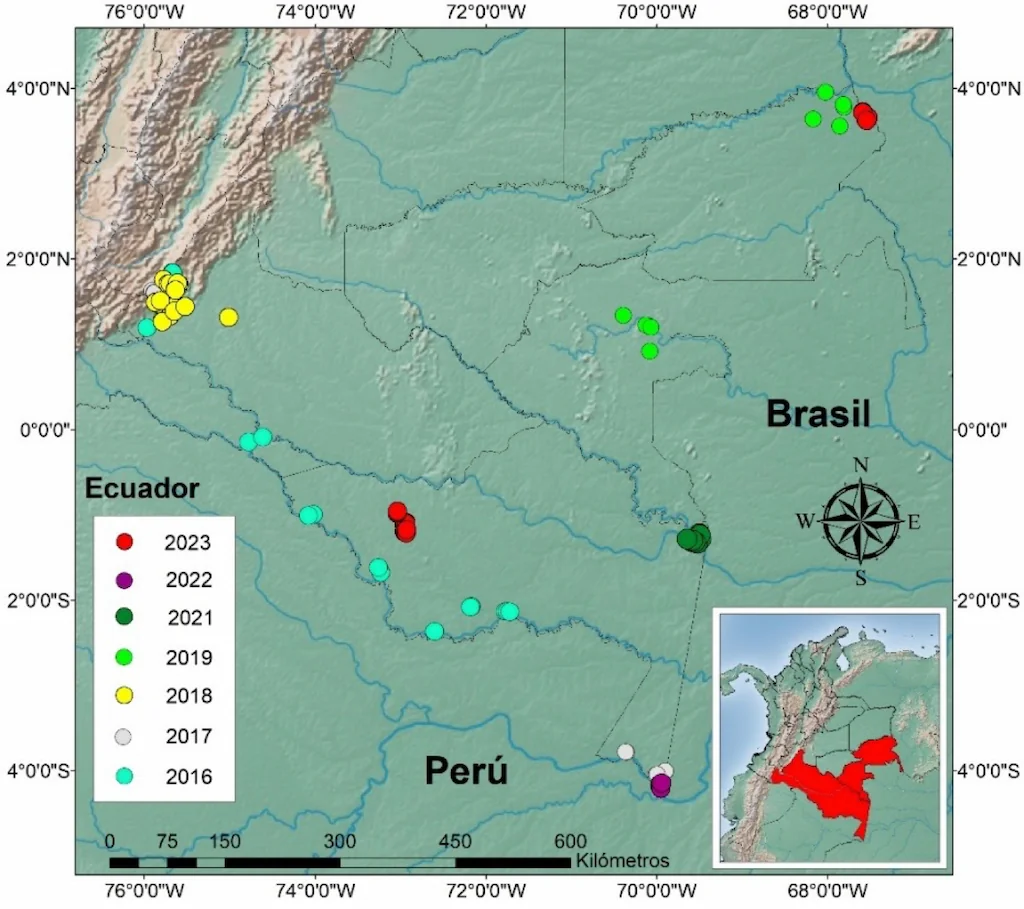
Figura 2. Mapa general de localidades muestreadas con TSBF a 2024. Fuente: Programa Ecosistemas y Recursos Naturales- Instituto SINCHI 2025.
En total, se identificaron 31 grupos taxonómicos. Las hormigas (Formicidae) y las termitas (Isoptera) fueron los grupos más abundantes, alcanzando las termitas densidades superiores a 10.000 individuos por metro cuadrado, lo que evidencia su rol estructural y funcional en los ecosistemas amazónicos. También destacaron por su abundancia las larvas de coleópteros, los dipluros, los coleópteros adultos, las arañas (Araneae), los diplópodos y los quilópodos (Figura 3). Estos grupos conforman el núcleo funcional de la macrofauna del suelo, al intervenir activamente en la fragmentación de la materia orgánica, la bioturbación, y la modulación de las condiciones físicas del suelo.
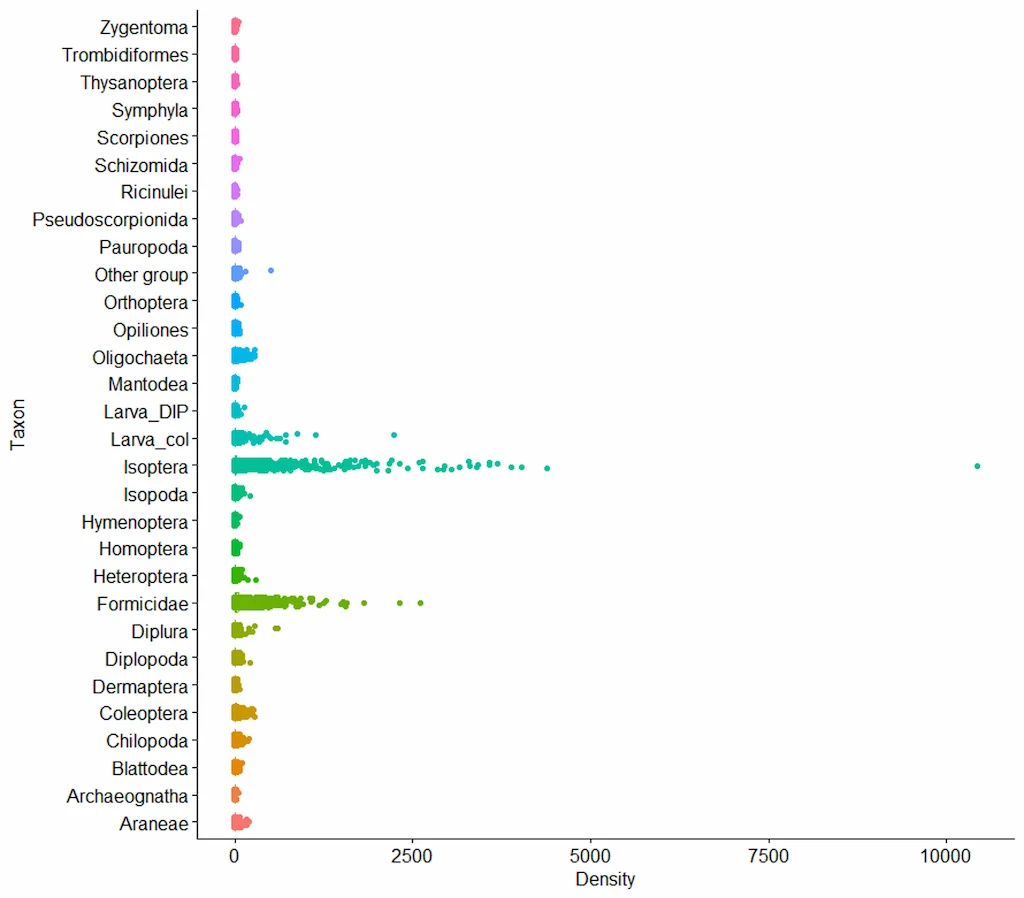
Figura 3. Densidad (ind/m2) de los grupos taxonómicos por monolitos en suelos amazónicos. Fuente: Programa Ecosistemas y Recursos Naturales- Instituto SINCHI 2025.
Los análisis de diversidad revelaron que en suelos de bosques primarios de tierra firme se pueden encontrar hasta 61 especies diferentes en un solo monolito (25 × 25 cm, 0–10 cm de profundidad). Aunque se observaron valores elevados de diversidad en pasturas y coberturas degradadas, estas fueron significativamente menores a las de bosques primarios y secundarios (Figura 4). En los bosques de várzea o rebalse, la diversidad también fue baja, probablemente debido a las condiciones anóxicas estacionales y la saturación de agua que restringen la presencia de especies menos tolerantes. Estos resultados muestran cómo las condiciones hidrológicas y el nivel de intervención antrópica determinan fuertemente la composición y riqueza faunística del suelo.
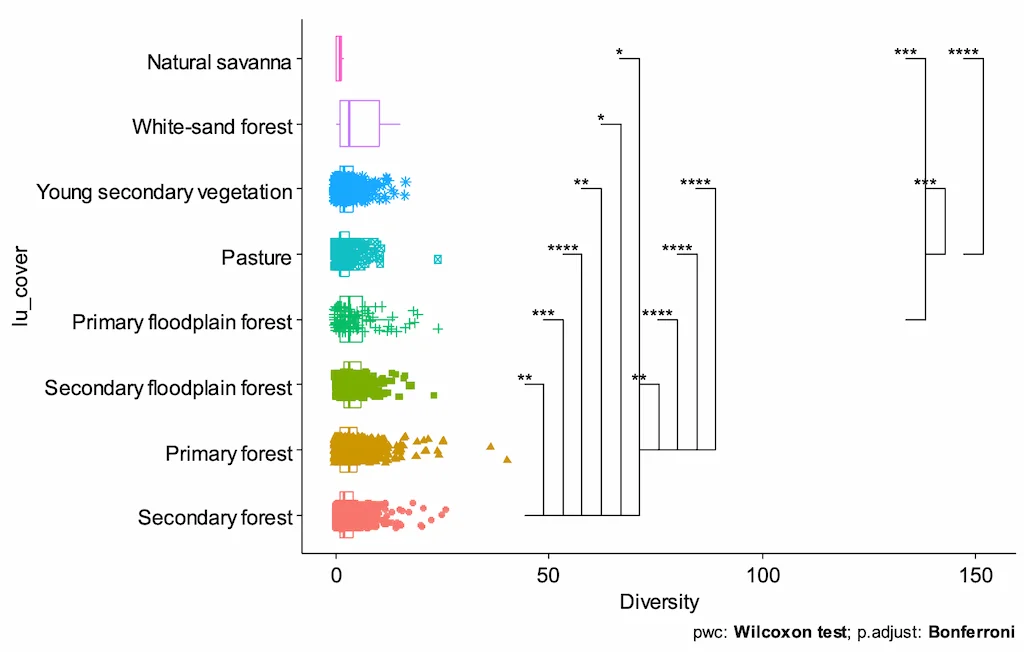
Figura 4. Diversidad por monolito de la macrofauna edáfica con respecto a las coberturas, barras muestras diferencias significativas de múltiples comparaciones por el método Benjamini & Hochberg. Fuente: Programa Ecosistemas y Recursos Naturales- Instituto SINCHI 2025.
En cuanto a la respuesta de los grupos taxonómicos frente a los distintos tipos de cobertura, se observó que las hormigas presentaron las mayores diferencias entre ambientes, seguidas de las lombrices, los coleópteros adultos y las termitas. La densidad general de la macrofauna fue significativamente distinta entre coberturas, lo que sugiere que cada componente responde de forma específica a variables como la calidad del suelo y la disponibilidad de recursos. A pesar de presentar densidades similares en algunas coberturas, la composición de grupos como coleópteros y oligoquetos varía marcadamente, indicando ensamblajes funcionales diferenciados. Estos resultados evidencian que no solo la abundancia, sino también la identidad funcional de los taxones es clave para entender los procesos ecológicos que regulan la biodiversidad edáfica.
La Figura 5 muestra los resultados del análisis de la densidad de los principales grupos taxonómicos de la macrofauna edáfica en distintos tipos de cobertura vegetal en la Amazonia colombiana. Este análisis se realizó a través de pruebas no paramétricas de Kruskal–Wallis para detectar diferencias globales entre coberturas, seguido de comparaciones post-hoc de Dunn con corrección de Benjamini & Hochberg, lo cual permite identificar qué coberturas difieren significativamente entre sí para cada grupo.
Cada subfigura dentro de la Figura 5 representa un grupo taxonómico clave: arañas, hormigas, termitas, coleópteros adultos, lombrices de tierra y la macrofauna total. En conjunto, estas comparaciones permiten evaluar cómo responde la densidad de cada grupo a distintos grados de perturbación del ecosistema, desde bosques primarios hasta coberturas altamente intervenidas como pasturas o zonas degradadas.
Las hormigas (Formicidae) fueron el grupo que mostró las diferencias más marcadas entre coberturas. Se observó un aumento significativo de su densidad en ambientes intervenidos, particularmente en pasturas, en comparación con bosques primarios y secundarios. Este patrón sugiere una alta tolerancia de las hormigas a la perturbación del hábitat y la expansión de especies generalistas o dominantes en condiciones alteradas. Su capacidad de adaptación a suelos compactados y su eficiencia para colonizar espacios abiertos podrían explicar su éxito en estos ambientes.
En contraste, las termitas (Isoptera) presentaron su mayor densidad en coberturas naturales, especialmente en bosques primarios, y una disminución progresiva en ambientes más perturbados. Este comportamiento indica que las termitas son más sensibles a cambios en la estructura del suelo y en la disponibilidad de materia orgánica lignificada, su principal fuente de alimento. Además, su dependencia de condiciones microclimáticas estables podría explicar su menor presencia en coberturas expuestas a mayor variabilidad térmica e hídrica.
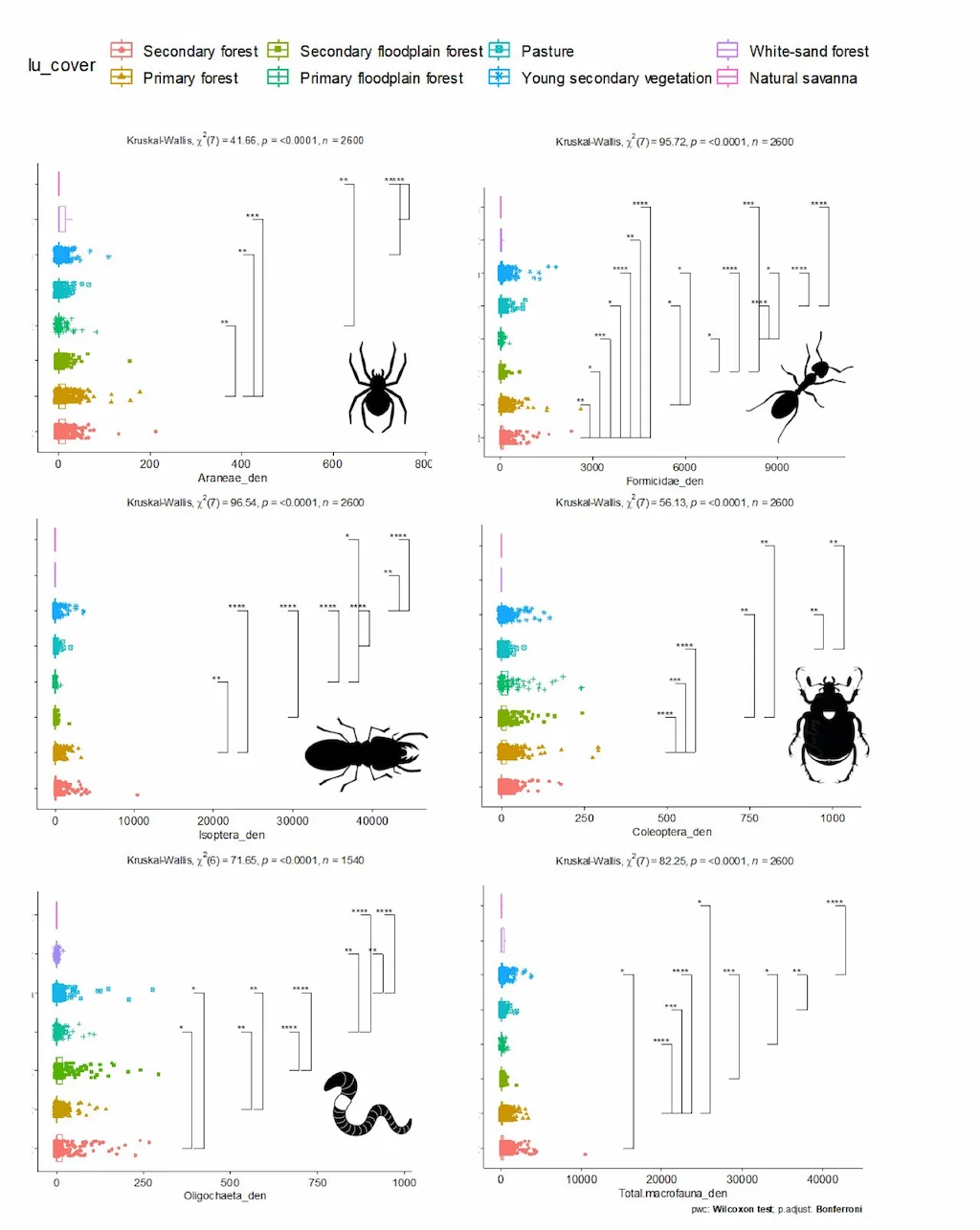
Figura 5. Kruskal–Wallis one-way y post-hoc Dunn test comparando la diversidad de a) arañas; b) hormigas, c) termitas, d) coleóptera adultos, e) lombrices y f) toda la macrofauna por coberturas. Barras muestras diferencias significativas de múltiples comparaciones por el método Benjamini & Hochberg. Fuente: Programa Ecosistemas y Recursos Naturales- Instituto SINCHI 2025.
Las lombrices de tierra (Oligochaeta) también presentaron una mayor densidad en coberturas naturales, aunque las diferencias no fueron tan pronunciadas como en el caso de las termitas. Su abundancia fue moderadamente mayor en bosques secundarios en comparación con pasturas y áreas degradadas. Dado su papel en la bioturbación y el ciclado de nutrientes, esta tendencia podría estar relacionada con la mayor disponibilidad de hojarasca y condiciones edáficas más estables en coberturas con mayor cobertura vegetal.
En el caso de los coleópteros adultos (Coleoptera) y las arañas (Araneae), las diferencias entre coberturas fueron menos marcadas, aunque se observan algunas tendencias. Los coleópteros adultos presentaron una ligera preferencia por bosques secundarios, mientras que las arañas mostraron una distribución relativamente homogénea, posiblemente influenciada por la estructura vegetal y la disponibilidad de refugios. En general, estos grupos parecen responder más a microhábitats locales que a las coberturas en sí mismas, aunque pueden ser útiles como indicadores complementarios.
Finalmente, al evaluar la macrofauna total, se observaron diferencias significativas entre coberturas, lo que sugiere que la comunidad en su conjunto responde de forma sensible a los cambios en el uso del suelo. Sin embargo, este resultado debe interpretarse con precaución, ya que valores elevados de densidad en algunas coberturas pueden estar determinados por el dominio numérico de uno o pocos grupos, como es el caso de las hormigas en pasturas. Por ello, es fundamental analizar tanto la densidad como la composición y diversidad de los grupos para obtener una visión más completa de la estructura funcional del suelo.
Esta Figura 5 evidencia que los distintos componentes de la macrofauna del suelo responden de manera diferenciada a la alteración de los ecosistemas. Algunos grupos como las hormigas aumentan su densidad en ambientes perturbados, mientras que otros como las termitas y lombrices se ven más favorecidos en condiciones de mayor integridad ecológica. Estas respuestas diferenciales resaltan la importancia de considerar múltiples grupos funcionales en el monitoreo de biodiversidad edáfica y en la evaluación del estado ecológico de los suelos amazónicos. Eso se amplía de gran manera en el reciente artículo científico publicado por el equipo de suelos del Instituto (Castro et al. 2025), en el que se demuestra que la recuperación de la macrofauna edáfica no es lineal ni homogénea, sino que depende del tiempo transcurrido desde el abandono, las condiciones locales del paisaje y las características físico-químicas del suelo.
El carbono orgánico del suelo (COS), como indicador clave de la calidad edáfica, mostró diferencias marcadas entre coberturas naturales e intervenidas. Se evidenció una fuerte correlación positiva entre el COS y la diversidad de la macrofauna, sugiriendo que suelos con mayor diversidad faunística tienden a tener mejores condiciones de fertilidad. Sin embargo, esta relación no fue evidente con la densidad, ya que ciertas especies dominantes en ambientes intervenidos presentaron altas abundancias, pero baja diversidad.
Los niveles de fósforo (P) y calcio (Ca) mostraron alta variabilidad entre sitios, a diferencia del magnesio (Mg), que fue más estable. El fósforo, en particular, fue clave en coberturas intervenidas, donde la macrofauna podría contribuir a su disponibilidad, favoreciendo procesos de regeneración edáfica. Esto resalta el papel de ciertos grupos de macrofauna como facilitadores de la recuperación de suelos degradados, al contribuir a la movilización y ciclado de nutrientes.
Recomendaciones y alternativas
Los resultados presentados confirman el papel clave de la biota del suelo en la sostenibilidad ecológica de los ecosistemas amazónicos, especialmente en suelos altamente meteorizados y de baja fertilidad. Las asociaciones micorrízicas arbusculares constituyen una estrategia fundamental para la nutrición vegetal, facilitando la absorción de fósforo y mejorando la tolerancia al estrés hídrico. Sin embargo, prácticas agrícolas como la quema de cobertura vegetal, el uso intensivo de herbicidas y la compactación del suelo comprometen gravemente estas asociaciones simbióticas. Se recomienda el desarrollo y la implementación de estrategias productivas que promuevan el uso sostenible del suelo, evitando el disturbio excesivo y fomentando condiciones que favorezcan la actividad micorrícica, como la cobertura permanente del suelo y la rotación diversificada de cultivos.
En cuanto a la macrofauna edáfica, los hallazgos evidencian que este componente sigue siendo poco explorado en la Amazonia colombiana, a pesar de su relevancia funcional. El aumento en la densidad de ciertos grupos en coberturas perturbadas contrasta con la disminución general de la diversidad, lo cual puede tener implicaciones negativas en el equilibrio ecológico del suelo. Por ello, es prioritario continuar con los inventarios, especialmente en departamentos y coberturas edáficas que hasta ahora han sido poco muestreadas. Se recomienda ampliar el uso de metodologías combinadas (TSBF, taxonomía molecular y morfológica) e incluir enfoques funcionales que permitan comprender con mayor precisión el papel ecológico de los diferentes grupos.
Asimismo, se requiere fortalecer los procesos de monitoreo a largo plazo e integrar los indicadores biológicos del suelo dentro de los sistemas nacionales de seguimiento ambiental y ordenamiento territorial, con el fin de vincular el conocimiento generado con las decisiones de manejo del suelo y los procesos de restauración ecológica.
Fichas del capítulo I


Fichas del capítulo I, 2024
Fichas del capítulo I, 2024

1. Avance en el conocimiento de la biodiversidad Amazónica 2024

2. Flora de la Amazonia colombiana

3. Nuevas especies de plantas para la ciencia en la flora amazónica colombiana

5. Fauna de la Amazonia colombiana
6. Microalgas de la cuenca del río Orito, Santuario Ingi-Ande

7. Evaluación microorganismos Metagenomica

8. Suelos de la Amazonia colombiana

9. Colección de no vasculares y líquenes
